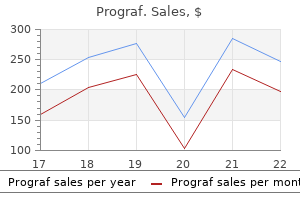
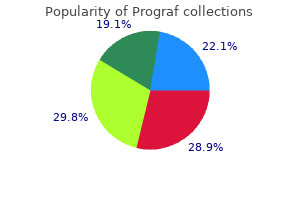
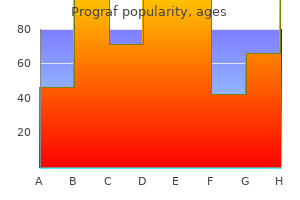
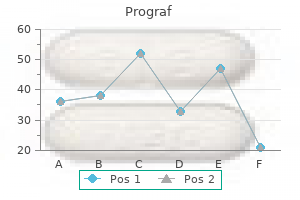
Prograf

Prograf
Prograf dosages: 5 mg, 1 mg
Prograf packs: 10 pills, 20 pills, 40 pills, 60 pills, 80 pills, 30 pills, 50 inhalers

Around 1 in 5 hiv infection from blood test 0.5 mg prograf generic with amex,000 infants is born with the condition and is often identified in fifty eight Brainstem Sensory vagal neurone Prevertebral sympathetic ganglion Spinal sensory neurone Spinal twine Intestinofugal neurone Neurocrine alerts: local and circulating Intrinsic sensory neurone Immune and tissue defence signals: native and systemic St re tch Gut lumen Signals from lumen antiviral brand names order prograf 0.5 mg line. Neurocrine indicators from enteric neuroendocrine cells and alerts from immune defence cells antiviral research conference generic 0.5mg prograf with mastercard. Some neuronal soma lie inside enteric ganglia within the gut wall; others have their our bodies in peripheral ganglia. Transduction varies with the modality of the stimulus, and normally causes depolarization of the receptor membrane (or hyperpolarization, in the retina). In mechanoreceptors, transduction might contain the deformation of membrane structure, which causes both strain or stretch-sensitive ion channels to open. Visual receptors share similarities with chemoreceptors: mild causes adjustments in receptor proteins, which activate G proteins, resulting in the launch of second messengers and altered membrane permeability. The quantitative responses of sensory endings to stimuli range tremendously and improve the flexibleness of the functional design of sensory techniques. Even unstimulated receptors show varying degrees of spontaneous background activity in opposition to which an increase or decrease in activity occurs with altering levels of stimulus. Though all receptors show these two phases, one or other might predominate, providing a distinction between quickly adapting endings that accurately report the speed of stimulus onset, and slowly adapting endings that sign the constant amplitude of a stimulus. Dynamic and static phases are mirrored within the amplitude and duration of the receptor potential and also within the frequency of motion potentials within the sensory fibres. Another broadly used classification divides receptors on the premise of their distribution within the body into exteroceptors, proprioceptors and interoceptors. Exteroceptors and proprioceptors are receptors of the somatic afferent components of the nervous system, whereas interoceptors are receptors of the visceral afferent pathways. Exteroceptors reply to external stimuli and are discovered at, or near, physique surfaces. They could be subdivided into the final or cutaneous sense organs and special sensory organs. General sensory receptors embrace free and encapsulated terminals in skin and near hairs; none of those has absolute specificity for a selected sensory modality. Special sensory organs are the olfactory, visible, acoustic, vestibular and taste receptors. Proprioceptors reply to stimuli to deeper tissues, especially of the locomotor system, and are concerned with detecting motion, mechanical stresses and place. They embrace Golgi tendon organs, muscle spindles, Pacinian corpuscles, other endings in joints, and vestibular receptors. Proprioceptors are stimulated by the contraction of muscular tissues, movements of joints and adjustments within the place of the body. They are important for the coordination of muscles, the grading of muscular contraction, and the maintenance of equilibrium. Interoceptors are discovered in the partitions of the viscera, glands and vessels, where their terminations embrace free nerve endings, encapsulated terminals and endings related to specialised epithelial cells. Free terminal arborizations occur within the endocardium, the endomysium of all muscular tissues, and connective tissue generally. Tension produced by excessive muscular contraction or by visceral distension often causes ache, notably in pathological states, which is incessantly poorly localized and of a deepseated nature. Polymodal nociceptors (irritant receptors) reply to a variety of stimuli such as noxious chemical compounds or damaging mechanical stimuli. They are mainly the free endings of fantastic, unmyelinated fibres that are broadly distributed in the epithelia of the alimentary and respiratory tracts; they may provoke protective reflexes. A variety of descriptions and terms have been utilized to cells of this technique in the older literature (see online text for details). This is an evolutionarily primitive arrangement, and the only examples remaining in humans are the sensory neurones of the olfactory epithelium. When activated, this type of receptor excites its neurone by neurotransmission across a synaptic gap. A neuronal receptor is a primary sensory neurone that has a soma in a craniospinal ganglion and a peripheral axon ending in a sensory terminal. All cutaneous sensors and proprioceptors are of this sort; their sensory terminals could additionally be encapsulated or linked to special mesodermal or ectodermal structures to form a part of the sensory apparatus. Many cells of the dispersed (or diffuse) neuroendocrine system are derived embryologically from the neural crest. Some � particularly, cells from the gastrointestinal system � at the second are recognized to be endodermal in origin. The traces below each kind of ending point out (top) their response (firing rate (vertical lines) and adaption with time) to an acceptable stimulus (below) of the duration indicated. They occur in all connective tissues, including those of the dermis, fasciae, capsules of organs, ligaments, tendons, adventitia of blood vessels, meninges, articular capsules, periosteum, perichondrium, Haversian techniques in bone, parietal peritoneum, partitions of viscera and the endomysium of all kinds of muscle. They also innervate the epithelium of the skin, cornea, buccal cavity, and the alimentary and respiratory tracts and their related glands. Within epithelia, free sensory endings lack Schwann cell ensheathment and are enveloped as a substitute by epithelial cells. Afferent fibres from free terminals may be myelinated or unmyelinated however are all the time of small diameter and low conduction velocity. When afferent axons are myelinated, their terminal arborizations lack a myelin sheath. In the dermis, they might be responsive to average cold or heat (thermoreceptors); light mechanical touch (mechanoreceptors); damaging warmth, cold or deformation (unimodal nociceptors); and damaging stimuli of several varieties (polymodal nociceptors). Similar fibres in deeper tissues may also sign extreme circumstances, which are experienced, as with all nociceptors, as ache or ache. Free endings in the cornea, dentine and periosteum could additionally be completely nociceptive. Special kinds of free ending are associated with epidermal structures within the pores and skin. They embrace terminals associated with hair follicles (peritrichial receptors), which branch from myelinated fibres within the deep dermal cutaneous plexus; the quantity, measurement and type of the endings are related to the dimensions and kind of hair follicle innervated. These endings respond mainly to motion when hair is deformed and belong to the rapidly adapting mechanoreceptor group. The cells are believed to be derived from the dermis, though a neural crest origin stays potential. They comprise many massive (50�100 nm) dense-core vesicles, presumably containing transmitters. Merkel endings are thought to be slow-adapting mechanoreceptors, aware of sustained pressure and delicate to the edges of utilized objects. They exhibit appreciable selection in their size, form and distribution however share a typical characteristic: particularly, that each axon terminal is encapsulated by non-excitable cells (Proske and Gandevia 2012). They are most concentrated in thick hairless pores and skin, particularly of the finger pads, where there may be as much as 24 corpuscles per cm2 in younger adults. Mature corpuscles are cylindrical in shape, roughly 80 �m long and 30 �m across, with their long axes perpendicular to the skin surface. They are oval, spherical or irregularly coiled and measure as much as 2 mm in size and 100�500 �m or extra across; the larger ones are seen to the bare eye. Each corpuscle has a capsule, an intermediate growth zone and a central core that contains an axon terminal. The capsule is formed by roughly 30 concentrically arranged lamellae of flat cells approximately 0. Adjacent cells overlap and successive lamellae are separated by an amorphous proteoglycan matrix that accommodates circularly oriented collagen fibres, intently utilized to the surfaces of the lamellar cells. The core consists of roughly 60 bilateral, compacted lamellae mendacity on each side of a central nerve terminal. Each corpuscle is equipped by a myelinated axon, which initially loses its myelin sheath and subsequently loses its ensheathing Schwann cell at its junction with the core. The naked axon runs by way of the central axis of the core and ends in a slightly expanded bulb. It is in contact with the innermost core lamellae, is transversely oval and sends brief projections of unknown function into clefts within the lamellae. The axon incorporates numerous large mitochondria, and minute vesicles, roughly 5 nm in diameter, which mixture reverse the clefts. The cells of the capsule and core lamellae are thought to be specialized fibroblasts however some could additionally be Schwann cells. Pacinian corpuscles are provided by capillaries that accompany the axon as it enters the capsule.
Quiescent osteoblasts and osteoprogenitor cells act because the principal reservoir of latest boneforming cells for remodelling or restore on the endosteal surfaces of resting grownup bone antiviral injection for chickenpox prograf 5 mg buy discount online. Bone endosteum is likely to hiv infection no ejaculation generic 1mg prograf with visa be essential in calcium homeostasis as a outcome of it provides a total surface space of roughly 7 hiv infection rates nyc prograf 5mg sale. It overlies the endosteal circumferential lamellae and encloses the medullary cavity. The vascular provide of a protracted bone depends on several points of influx that feed complex and regionally variable sinusoidal networks inside the bone. One or two primary diaphysial nutrient arteries enter the shaft ob liquely through nutrient foramina, which lead into nutrient canals. Their websites of entry and angulation are nearly fixed and characteristi cally directed away from the dominant growing epiphysis. Osteoprogenitor cells, osteoblasts, osteoclasts and other cells necessary in the turnover and homeostasis of bone tissue lie in these layers. Concentric lamellae encompass the central Haversian canal of each complete osteon; they comprise the darkish lacunae of osteocytes and the canaliculi, which are occupied in life by their dendrites. B, A high-power view of osteocytes within lamellae; a Haversian canal is seen on the best. A, Irregular trabeculae of bone, surrounded by bone marrow haemopoietic and adipose tissue (haematoxylin and eosin stain). B, the same subject seen under polarized mild, demonstrating lamellar, non-osteonic bone with lamellae oriented in several directions in different areas. The endosteal vessels are vulnerable throughout surgical operations, similar to intramedullary nailing, which contain passing metallic implants into the medullary canal. The former are direct branches of neighbouring systemic vessels; the latter come from periarticular vas cular arcades formed on nonarticular bone surfaces. Numerous vascu lar foramina penetrate bones close to their ends, often at pretty particular sites; some are occupied by arteries but most include thinwalled veins. Within bone, the arteries are uncommon in consisting of endothelium with solely a skinny layer of supportive connective tissue. The epiphysial and metaphysial arterial provide is richer than the diaphysial provide. Medullary arteries within the shaft give off centripetal branches, which feed a hexagonal mesh of medullary sinusoids that drain into a large, thinwalled central venous sinus. They additionally possess cortical branches, which pass via endosteal canals to feed fenestrated capillaries in Haversian systems. The central sinus drains into veins that retrace the paths of nutrient arteries, sometimes piercing the shaft elsewhere as unbiased emissary veins. A capillary community permeates the deeper, osteogenic perios teum from this exterior plexus. At muscular attachments, periosteal and muscular plexuses are confluent and the cortical capillaries then drain into interfascicular venules. The giant nutrient arteries of epiphyses form many intraos seous anastomoses, their branches passing in the path of the articular surfaces throughout the trabecular areas of the bone. The latter often pierce the thin hypochondral compact bone to enter, and typically traverse, the calcified zone of articular cartilage, earlier than returning to the epiphy sial venous sinusoids. In immature lengthy bones, the availability is analogous however the epiphysis is a discrete vascular zone. Epiphysial and metaphysial arteries enter on each side of the expansion cartilage and barely, if ever, anastomose. Note the contrasting supplies of the diaphysis, metaphysis and epiphysis, and their connections with periosteal, endosteal, muscular and peri-articular vessels. The marrow cavity accommodates a large central venous sinus, a dense community of medullary sinusoids, and longitudinal medullary arteries and their circumferential rami. Growth cartilages are probably equipped from each sources, and from an anastomotic collar within the adjoining periosteum. Metaphysial bone is nourished by terminal branches of metaphysial arteries and by main nutrient arteries of the shaft, which kind terminal blindended sprouts or sinusoidal loops within the zone of advancing ossification. Young peri osteum is more vascular; its vessels talk more freely with these of the shaft than their grownup counterparts and provides off extra metaphysial branches. Large, irregular bones such because the scapula and innominate not only obtain a periosteal provide however are also often provided by massive nutrient arteries that penetrate directly into their cancellous bone, the 2 methods anastomosing freely. Short bones receive numerous fine vessels that provide their compact and cancellous bone and medullary cavities from the periosteum. Flatter cranial bones are provided by numerous periosteal or mucoperiosteal vessels. Osteoblasts possess receptors for several neuropeptides found in these nerves, together with neuropeptide Y, calcitonin generelated peptide, vasoactive intestinal peptide and substance P, indicating that bone has a posh autonomic and sensory innervation. Most bones are fashioned by a means of endochondral ossi fication, by which preformed cartilage templates (models) define their preliminary shapes and positions, and the cartilage is replaced by bone in an ordered sequence. At centres of ossification, mesenchymal stem cells differentiate into osteoprogenitor cells, which proliferate around the branches of a capillary community, forming incomplete layers of osteoblasts in touch with the primitive bone matrix. The cells are polarized, and secrete osteoid only from the floor that faces away from the blood vessels. The earliest crystals appear in affiliation with Innervation Nerves are most quite a few in the articular extremities of lengthy bones, vertebrae and larger flat bones, and in periosteum. The cartilage cells within the shaft have hypertrophied and this area is surrounded by a fragile tube or collar of subperiosteal bone (red). Crystal forma tion subsequently extends into collagen fibrils within the surrounding matrix, producing an early labyrinth of woven bone, the first spon giosa. As layers of calcifying matrix are added to the early trabeculae, osteoblasts become enclosed inside primitive lacunae. These new osteocytes retain intercellular contact via their fine cytoplasmic processes (dendrites) and, as these elongate, matrix condenses round them to type canaliculi. As matrix secretion and calcification proceed, trabeculae thicken and vascular areas turn into narrower. Where bone remains trabecular, the process slows and the areas between trabeculae turn into occupied by haemopoietic tissue. Where compact bone is forming, trabeculae con tinue to thicken and vascular spaces proceed to slim. Meanwhile, the collagen fibres of the matrix, secreted on the partitions of the narrowing spaces between trabeculae, become organized as parallel, longitudinal or spiral bundles, and the cells they enclose occupy concentric sequen tial rows. These irregular, interconnected plenty of compact bone every have a central canal and are known as main osteons (primary Haversian systems). They are later eroded, together with the intervening woven bone, and changed by generations of mature (secondary) osteons. While these adjustments are occurring, mesenchyme condenses on the outer surface to form a fibrovascular periosteum. Bone is laid down increasingly by new osteoblasts, which differentiate from osteoprogeni tor cells in the deeper layers of the periosteum. Modelling of the growing bone is achieved by various charges of resorption and deposition at different sites. This low-magnification micrograph exhibits the first ossification centre in a human fetal bone. Abbreviations; G, development zone; H, hypertrophic zone; O, ossification zone; R, remodelling zone. Endochondral ossification the hyaline cartilage mannequin that forms during embryogenesis is a mini ature template of the bone that will subsequently develop. Matrix vesicles originat 92 ing from chondrocytes within the proliferation zone are most evident within the intercolumnar areas, where they seem to provoke crystal formation. At the identical time, cells within the deep layer of perichondrium around the centre of the cartilage mannequin differentiate into osteoblasts and form a peripheral layer of bone. The periosteal collar, which overlies the calcified cartilaginous partitions of degenerate chondrocyte lacunae, is invaded from the deep layers of the periosteum (formerly perichondrium) by osteogenic buds. These are blindended capillary sprouts which are accompanied by osteopro genitor cells and osteoclasts. Lacunae are enlarged, and matrix partitions are decreased in width and exhibit elevated staining density following cartilage calcification. Spicules of cartilaginous remnants (pale blue) serve as surfaces for the deposition of osteoid (dark blue), shown in the upper half of the sector.
Cellular improvement of the cerebellum the cerebellum consists of a cortex early stages of hiv infection symptoms discount prograf 0.5mg without a prescription, beneath which are buried a collection of deep nuclei symptoms untreated hiv infection prograf 0.5 mg purchase without prescription. Thus hiv infection and pregnancy purchase prograf 1 mg without prescription, the cerebellum has an intraventricular portion (cells proliferating from the ventricular zone) and an extraventricular portion (cells proliferating from the external germinative layer) throughout development. The extraventricular portion turns into larger on the expense of the intraventricular part, the so-called extroversion of the cerebellum. Before the top of the third month, the primary mass of the cerebellum is extraventricular. The developed cerebellar cortex contains three layers: namely, the molecular layer, the Purkinje layer and the granular layer. The radial glia play a role in guiding the Purkinje cells to the meningeal surface of the cerebellar anlage. During this early stage of cerebellar improvement, which is dominated by the manufacturing and migration of efferent cerebellar neurones, the surface of the cerebellar anlage stays easy. The extroversion of the cerebellum begins later when cells of the external germinative layer, additionally termed the superficial matrix, begin proliferation and migration. These cells produce the granule cells, which migrate inwards along the radial glia, through the layers of Purkinje cells, settling deep to them in the granular layer. Proliferation and migration of granule cells lead to a great rostrocaudal enlargement of the meningeal surface of the cerebellum, forming the transverse fissures and transforming the multicellular layer of Purkinje cells right into a monolayer. Purkinje cells and nuclear cells are fashioned previous to the granule cells, and granule cells function the recipient of the main afferent (mossy fibre) system of the cerebellum. Thus, the development of the efferent neurones of the cerebellar cortex and nuclei precedes the event of its afferent group. The early bilateral cerebellar anlage is modified right into a unitary structure by fusion of the bilateral intraventricular bulges and the disappearance of the ependyma at this site, the merging of the left and right primitive cerebellar cortex over the midline, and the event of the cerebellar commissure by ingrowth of afferent fibres and outgrowth of efferent axons of the medial cerebellar nucleus. These Purkinje cells will grow axons that connect to neurones within the vestibular nuclei and the fastigial nucleus. The lateral clusters belong to the lengthy run hemispheres and will develop axons terminating within the interposed and dentate nuclei. The sharp border within the efferent projections from the vermis and hemispheres is thus established at an early age. These clusters will give rise to Purkinje cell zones in the grownup cerebellum, which project to a single vestibular or cerebellar nucleus. In the developing human mind, only the external germinative layer could be seen at 17�18 weeks; the Purkinje cells become obvious between 20 and 23 weeks. After 30 weeks, four layers can be acknowledged: the external germinative layer (external granular layer), fashioned by 6�8 rows of densely packed small round cells; the Purkinje cell layer, shaped by 5�6 layers of bigger, spherical, immature neurones external to the internal granular layer; and the molecular layer, containing cells that resemble exterior granular layer cells in migration (Lavezzi et al 2006). From 5 to 7 months, the Purkinje cells are decreased in number and extra widely spaced, displaying mature polygonal somata with evident axons and dendrites. Relatively little is understood about the molecular mechanisms involved in cerebellar growth in the human (but see Manto and Jissendi (2012), Yu et al (2013), Martinez et al (2013)). Very small preterm infants are at explicit threat of later cerebellar dysfunction with penalties for cognition and behavior. A, Purkinje cells and cells of the cerebellar nuclei are produced by the ventricular zone and migrate to their future positions. The cells of the superficial matrix (the exterior granular layer) take their origin from the ventricular epithelium on the caudal pole of the cerebellar anlage and migrate rostrally over its surface. B, After migration, the Purkinje cells constitute a multicellular layer beneath the external germinative layer. C, Granule cells produced by the external germinative layer migrate inwards through the Purkinje cell layer to their place in the inner granular layer. The Purkinje cells are located in 5 multicellular clusters (stars) on each side of the midline. The anlage of the dentate nucleus occupies the centre of probably the most lateral Purkinje cell cluster. The cells of the dorsal part of the alar (dorsolateral) plates proliferate and invade the roof plate, which due to this fact thickens and is later divided into corpora bigemina by a median groove. Caudally, this groove turns into a median ridge, which persists within the adult because the frenulum veli. The corpora bigemina are later subdivided into the superior and inferior colliculi by a transverse furrow. The purple nucleus, substantia nigra and reticular nuclei of the midbrain tegmentum may first be outlined on the end of the third month. A high degree of geometric order exists within the growing retinotectal projection (the equal of the retinogeniculate projection) and in the tectospinal projection. In the absolutely developed mind, it persists as a slight groove extending from the interventricular foramen to the cerebral aqueduct. The thin roof plate remains epithelial but invaginated by vascular mesenchyme, the tela choroidea of the choroid plexuses of the third ventricle. Later, the lateral margins of the tela endure an identical invagination into the medial partitions of the cerebral hemispheres. The flooring plate thickens because the nuclear masses of the hypothalamus and subthalamus develop. Later, its cavity turns into relatively decreased in diameter and, within the grownup mind, it varieties the cerebral aqueduct. The basal (ventrolateral) plate of the midbrain increases in thickness to type the cerebral peduncles, that are at first of small size but enlarge quickly after the fourth month, when their numerous fibre tracts begin to appear in the marginal zone. The neuroblasts of the basal plate of mesencephalon 2 give rise to the nuclei of the oculomotor nerve and a few grey lots of the tegmentum, while the nucleus of the trochlear nerve stays within the region of the isthmus rhombencephali. At this early time, two eye fields are separated by the future neurohypophysis within the floor of the longer term D1. After head folding, the attention fields broaden as two lateral optic evaginations, which turn out to be optic vesicles, one on all sides of the early mind. For a time, they impart with the cavity of the prosencephalon by comparatively broad openings. The distal parts of the optic vesicles broaden, whereas the proximal elements turn out to be the tubular optic stalks. The optic chiasma is usually thought to be the boundary between diencephalon and telencephalon. As probably the most rostral portion of the prosencephalon enlarges, it curves ventrally, and two additional diverticula increase quickly from it, one on each side. These diverticula, that are rostrolateral to the optic stalks, subsequently form the cerebral hemispheres. Their cavities are the rudiments of the lateral ventricles and so they communicate with the median part of the forebrain cavity by comparatively wide openings, which ultimately turn out to be the interventricular foramina. The rostral part of the forebrain, including the rudiments of the cerebral hemispheres, consists of the telencephalon and the caudal part of the diencephalon; each contribute to the formation of the third ventricle, although the latter predominates. Diencephalon the diencephalon, D2, is broadly divided by the hypothalamic sulcus into dorsal (pars dorsalis diencephali) and ventral (pars ventralis diencephali) components; every contributes to diverse neural structures. Caudal to the thalamus, the lateral and medial geniculate our bodies, or metathalamus, are recognizable at first as surface depressions on the inner aspect and as elevations on the exterior facet of the lateral wall. As the thalami enlarge as easy ovoid masses, the wide interval between them gradually narrows into a vertically compressed cavity that types the larger a half of the third ventricle. After a time, these medial surfaces could come into contact and become adherent over a variable space, the connection (single or multiple) constituting the interthalamic adhesion or massa intermedia. The caudal growth of the thalamus excludes the geniculate bodies from the lateral wall of the third ventricle. Later, with the event of the projection fibres (corticofugal and corticopetal) of the neocortex, the thalamus turns into related to the inner capsule, which intervenes between it and the lateral part of the corpus striatum (lentiform nucleus). Ventral to the hypothalamic sulcus, the lateral wall of the diencephalon, in addition to median derivatives of its ground plate, varieties a large part of the hypothalamus and subthalamus. The epithalamus, which incorporates the pineal gland, the posterior and habenular commissures and the trigonum habenulae, develops in affiliation with the caudal part of the roof plate and the adjoining areas of the lateral walls of the diencephalon. At an early interval (12�20 mm crown�rump length), the epithalamus in the lateral wall tasks into the third ventricle as a easy ellipsoid mass, larger than the adjoining mass of the (dorsal) thalamus and separated from it by a well-defined epithalamic sulcus. In subsequent months, progress of the thalamus rapidly overtakes that of the epithalamus and the intervening sulcus is obliterated. Thus, structures of epithalamic origin ultimately are relatively diminutive topographically. The pineal gland arises as a hollow outgrowth from the roof plate, instantly adjoining the mesencephalon. Its distal part turns into strong by cellular proliferation but its proximal stalk remains hole, containing the pineal recess of the third ventricle.
In females hiv infection rate minnesota buy prograf 5 mg lowest price, oestrogens are probably to describe the hiv infection cycle 5 mg prograf generic visa keep vellus hairs; postmenopausal reduction of oestrogens may permit stronger facial and bodily hair growth hiv infection rates florida cheap 0.5mg prograf mastercard. When first shaped, sebum is a fancy combination of over 50% di- and triglycerides, with smaller proportions of wax esters, squalene, ldl cholesterol esters, cholesterol and free fatty acids. At start, sebaceous glands are quite giant, regressing later until stimulated once more at puberty. At that point, sebaceous gland progress and secretory activity increase tremendously in both men and women, beneath the influence of androgens (testicular and suprarenal), which act immediately on the gland. Excessive quantities of sebum may become impacted within the duct and this, related to hyperkeratinization, might result in blockage and formation of a comedone. Oestrogens have an effect reverse to that of androgens, and sebum secretion is significantly lower in ladies, becoming tremendously decreased after the age of 50 years. The secretory area may be as much as 2 mm extensive and its coils typically anastomose to type a labyrinthine network. Each coil is lined by cuboidal secretory cells whose apical cytoplasm tasks into the lumen and basally is involved with a layer of myoepithelial cells within a thick basal lamina. The secretory cells include vacuoles, vesicles and dense granules of various size and inner construction; the numbers and character vary with the cycle of synthesis and discharge. It is managed by adrenergic nerves and is delicate to adrenaline (epinephrine) and noradrenaline (norepinephrine). The secretion is initially sterile and odourless, nevertheless it undergoes bacterial decomposition to generate odorous and musky compounds, together with short-chain fatty acids, and steroids such as 5-androstenone. In many animals these are potent pheromonal alerts but their position in humans is less certain. Obstruction of apocrine sweat ducts and related higher hair follicles in the axillae, breast areolae and pubic region, primarily in girls, is assumed to underlie Fox�Fordyce disease. Chronic inflammation involving the skin bearing apocrine glands results in a painful, sometimes debilitating condition referred to as hidradenitis suppurativa. They present the standard options of smooth muscle cells and are separated by slim areas containing collagen fibres and unmyelinated noradrenergic sympathetic axons. The muscles are connected to the bulge region of the follicles by elastin fibrils, and are directed obliquely and towards the side to which the hair slopes superficially. The sebaceous gland occupies the angle between the muscle and the hair follicle, and muscle contraction helps to expel the gland contents. Contraction tends to pull the hair right into a more vertical position and to elevate the epidermis surrounding it into a small hillock (goose bump), dimpling the skin floor the place the muscle is inserted superficially. Arrector pili muscles are absent from facial, axillary and pubic hairs, eyelashes and eyebrows, and from the hairs around the nostrils and the exterior auditory meati. In thick hairless skin, sweat glands discharge alongside the centres of friction ridges, incidentally offering fingerprint patterns for forensic evaluation. Sweat glands have an necessary thermoregulatory function, and their secretion enhances grip and sensitivity of the palms and soles. Eccrine sweat glands are absent from the tympanic membrane, margins of the lips, nail bed, nipple, internal preputial surface, labia minora, penis and clitoris, where apocrine glands are located. They are numerous elsewhere, their frequency ranging from 80 to over 600/cm2, depending on position and genetic variation. There are many sweat glands on the face and flexor elements of the arms, and least on the surfaces of the limbs. People indigenous to hotter climates are inclined to have more sweat glands than these indigenous to cooler areas. The glands Microscopically, the secretory coil consists of a pseudostratified epithelium enclosing a lumen. Three kinds of cell have been described: clear cells, from which most of the secretion is derived; dark cells, which share the identical lumen; and myoepithelial cells. Clear cells are roughly pyramidal in form, and their bases relaxation on the basal lamina or are involved with the myoepithelial cells. Their apical plasma membranes line lateral intercellular canaliculi, which connect with the primary lumen. The basolateral plasma membranes are highly folded, interdigitating with apposed clear cells, they usually have the basal membrane infoldings typical of cells involved in transcytotic fluid and ion transport from the interstitial fluid to the glandular lumen. Their cytoplasm accommodates glycogen granules, mitochondria, tough endoplasmic reticulum and a small Golgi advanced, but few different organelles. Dark cells are pyramidal and lie closer to the lumen, such that their broad ends form its lining. Their cytoplasm accommodates a well-developed Golgi advanced, numerous vacuoles and vesicles, and dense glycoprotein granules, which they secrete by a typical merocrine mechanism. The intradermal sweat duct consists of an outer basal layer and an inner layer of luminal cells connected by quite a few desmosomes. The intraepidermal sweat duct (acrosyringium) is coiled and consists of two layers of cells, which, developmentally, are different from the encompassing keratinocytes and may be distinguished from them by the presence of keratin K19. The outer cells close to the surface contain keratohyalin granules and lamellar granules, and undergo typical cornification. The internal cells, from a mid-epidermal level, comprise numerous vesicles, endure an incomplete type of cornification, and are largely shed into the lumen on the degree of the cornified epidermal layer. Sweat is a transparent, odourless fluid, hypotonic to tissue fluid, and accommodates primarily sodium and chloride ions, but in addition potassium, bicarbonate, calcium, urea, lactate, amino acids, immunoglobulins and different proteins. It is modified as it passes alongside the duct by the motion mainly of the basal cells, which resorb sodium, chloride and a few water. The sweat glands are capable of producing up to 10 litres of sweat per day, in response to thermal, emotional and taste stimuli, mediated by unmyelinated sympathetic cholinergic fibres. Thermoregulation is coordinated via a heat centre in the preoptic area, in and close to the rostral hypothalamus, which reacts to modifications in blood temperature and afferent stimuli from the pores and skin by initiating applicable responses similar to controlling cutaneous blood provide, the speed and volume of sweat secretion (for evaporation at the surface) and shivering. They encompass compacted, anucleate, keratin-filled squames in two or three horizontal layers. Ultrastructurally, the squames include carefully packed filaments that lie transversely to the course of proximodistal development, and are embedded in a dense protein matrix. The hardness of nail is determined by the arrangement and cohesion of the layers of squames and their internal fibres, and never by the calcium content material. The water content material of nail is low but nail is 10 occasions extra permeable to water than the general epidermis. The softness and elasticity of the nail plate are related to its diploma of hydration. The nail apparatus consists of the nail plate, proximal and lateral nail folds, nail matrix, nail mattress and hyponychium. The nail plate arises from compacted cornified epithelial cells derived from the dorsal, intermediate and ventral nail matrices. It is densely adherent to the matrices on its undersurface however turns into a free construction distal to the onychodermal band, the place it separates from the nail mattress. The dorsal aspect of the nail plate originates from the extra proximal regions of the germinal matrix, i. It is roughly rectangular in shape and is generally convex in each longitudinal and transverse axes. The surface of the nail plate might present nice longitudinal ridges, and its undersurface is grooved by corresponding ridges within the nail bed. Disturbances of growth sample or illness might lead to transverse ridging or grooves. They are bounded by the attachment of the pores and skin to the lateral facet of the distal phalanx margin and the lateral nail. The proximal nail fold provides the seen proximal border to the nail apparatus. The epidermis of the superficial layer lacks hair follicles and epidermal ridges, and its cornified distal margin extends over the nail plate as the cuticle or eponychium. It adheres to the dorsal side of the nail plate and overlies the foundation of the nail. Nail matrix the nail matrix is the principle supply of the nail plate and it can be subdivided into three elements. Proximally, the dorsal matrix is outlined because the volar surface (undersurface) of the proximal nail fold. The intermediate matrix (germinal matrix) starts the place the dorsal matrix folds again on itself and extends so far as the distal portion of the lunula. The ventral matrix (sterile matrix) is the remainder of the nail bed; it starts at the distal border of the lunula and ends on the hyponychium.
Effectively hiv infection rates wikipedia prograf 1mg proven, water is held in place by proteoglycans hiv infection rate thailand 5 mg prograf purchase overnight delivery, which are themselves held in place by the collagen community antivirus windows xp cheap prograf 1 mg without a prescription. Other constituents of cartilage embody dissolved salts, noncollagenous proteins, and glycoproteins. Proteoglycan complexes bind to the floor of these fibres by way of their monomeric side chains and hyperlink them collectively. The arrangement of glycosaminoglycans and core protein of the proteoglycan monomer is illustrated in the growth. A, A low-power view of human rib, showing perichondrium (P), young chondroblasts (Cb) embedded in pale-staining interterritorial matrix, and mature chondrocytes (Cc) embedded within the basophilic interterritorial matrix (centre and right). B, Higher magnification of hyaline cartilage in human bronchial wall, showing isogenous teams of chondrocytes (C). Note the more deeply stained basophilic zones (B) (rich in acidic proteoglycans) across the cell clusters, with older, paler-staining matrix (M) between clusters. The central cell has an lively euchromatic nucleus with a distinguished nucleolus, and its cytoplasm accommodates concentric cisternae of tough endoplasmic reticulum, scattered mitochondria, lysosomes and glycogen aggregates. The plasma membrane bears numerous quick filopodia which project into the encompassing matrix. The latter shows a delicate feltwork of collagen fibrils within finely granular interfibrillary material. No pericellular lacuna is current; the matrix separates the central chondroblast from the cytoplasm of two adjoining chondroblasts (left, and crescentic profile). Chondrocytes synthesize and secrete all the main elements of the cartilage matrix, and their ultrastructure is typical of cells that are lively in making and secreting proteins. The nucleus is round or oval, appears euchromatic and possesses one or more nucleoli. The cyto plasm is filled with tough endoplasmic reticulum, transport vesicles and Golgi complexes, and contains many mitochondria and frequent lyso somes, along with numerous glycogen granules, intermediate fila ments (vimentin) and pigment granules. When these cells mature to the comparatively inactive chondrocyte stage, the nucleus turns into hetero chromatic, the nucleolus smaller, and the protein artificial machinery a lot lowered; the cells may accumulate giant lipid droplets. Polypeptide chains are assembled into triple helices, and a few carbohydrate is added. After transport to the Golgi equipment, the place further glycosylation happens, the resulting procollagen molecules are secreted into the extracellular area. Termi nal registration peptides are cleaved from their ends, forming tropocol lagen molecules, and the final meeting into collagen fibrils takes place. Matrix turnover is way slower in cartilage than in more metaboli cally active tissues. Collagen turnover is particularly slow, leaving it susceptible to the slow strategy of nonenzymatic glycation, which makes the tissue yellow, stiff and weak to damage (DeGroot et al 2004). Proteoglycans are turned over faster than collagen, with an esti mated turnover time of 5 years for adult humans. Certainly, the ability of the matrix to deform under load makes it difficult for hole blood vessels to persist within the tissue past early childhood, however a limited vascular provide is commonly discovered on the cartilage floor, from the place it could revas cularize the tissue following damage or degeneration. Metabolite transport severely limits cell density and metabolic price in the adult, and this in flip restricts carti lage thickness to a number of millimetres (Junger et al 2009). In the bigger cartilages, and during the speedy development of some fetal cartilages, vascular cartilage canals penetrate the tissue at intervals, providing an extra source of vitamins. In some circumstances these canals are short-term buildings but others persist all through life. Hyaline cartilage Hyaline (glassy) cartilage has a homogeneous, opalescent look, sometimes appearing bluish. Within such a chondron, daughter cells of a typical chondroblast typically meet at a straight line. The pericellular matrix closest to the cells is usually lacking in collagen fibrils, however rich in proteoglycans that may exhibit basophilic and metachromatic staining. Three-dimensional digital volumetric fluorescence imaging of serially sectioned, eosin-Y and acridine orange-stained tissue. Note the adjustments in dimension and spatial distribution of articular chondrocytes via the thickness of the cartilage. Schumacher, Department of Bioengineering and Whitaker Institute of Biomedical Engineering, University of California, San Diego. After adolescence, hyaline cartilage may turn out to be calcified as part of the conventional means of bone development, or as an agerelated, degen erative change. In costal cartilage, the matrix tends to fibrous striation, especially in old age when cellularity diminishes. The xiphoid course of and the cartilages of the nose, larynx and trachea (excepting the elastic cartilaginous epiglottis and corniculate cartilages) resemble costal automobile tilage in microstructure. Hyaline cartilage is the prototypical form, but it varies more with age and location than both elastic or fibrocartilage. The thickness of articular cartilage varies from 1 to 7 mm (typically 2 mm) in different joints, and decreases from center to old age. Central regions are inclined to be thickest on convex osseous surfaces, and thinnest on concave surfaces. Articular cartilage is usually too skinny and stiff to be a great shock absorber, though shock absorption may be vital the place there are multiple cartilagecovered surfaces, as in the carpus and tarsus. Embedded in the deep calcified zone, fibrils rise vertically by way of the radial zone towards the cartilage surface, where they seem to reorientate to run parallel to the floor in the tangential zone. Their threedimensional orientation can be appreciated by repeatedly piercing the cartilage floor with a needle; this creates a series of permanent elongated splits in the surface, which can be stained by Indian ink. The ensuing cut up line sample (Meachim et al 1974) reveals the predominant instructions of collagen bundles in the cartilage tangential zone, which can be related to inner traces of pressure gener ated during joint movement. The tangential (or superficial) zone has relatively small, elongated cells oriented parallel to the floor. Here, the chondrocytes are large and rounded, and surrounded by collagen fibrils in a spread of indirect ori entations. Deeper still, in the radial zone, the cells are sometimes disposed in vertical columns, interspersed with vertical collagen fibrils. An undulating band often identified as the tidemark indicates the beginning of the deepest zone, the zone of calcified cartilage, which has mechanical properties intermediate between cartilage and bone. This calcified zone is keyed into the subchondral bone by fine ridges and interdigitations, which serve to prevent shearing (gliding) movements between cartilage and bone. With age, articular cartilage thins by upward advancement of the tidemark, and gradual replacement of calcified cartilage by bone. Superficial cells are misplaced progressively from regular younger joint surfaces, to be replaced by cells from deeper layers. Agerelated reductions in cell quantity and activity, and biochemical adjustments in the extracellular matrix, significantly affect the superficial zone of articular cartilage, growing the danger of mechanical failure and of osteoarthritis (Lotz and Loeser 2012). Articular cartilage derives vitamins by diffusion from vessels of the synovial membrane, synovial fluid and hypochondral vessels of an adjacent medullary cavity, some capillaries from which penetrate and infrequently traverse the calcified cartilage zone. Small molecules freely traverse articular cartilage, with diffusion coefficients about half these in aqueous resolution. Larger molecules have diffusion coefficients inversely related to their molecular dimension. In each instances, chondrocytes undergo a sequence of cell divisions and hypertrophy (with cells forming into columns) followed by cell dying, and ossification by invading osteoblasts. Fibrocartilage Fibrocartilage is a dense, whitish tissue with a distinct fibrous texture. It forms the intervertebral discs of the backbone and menisci of the knee, in addition to smaller constructions such because the glenoid and acetabular labra, and the liner of bony grooves for tendons. Chondroblasts lie between coarse collagen sort I fibres (blue) derived from the anulus fibrosus. Histologically, fibrocartilage is intermediate between dense fibrous connective tissues such as tendon and ligaments, and hyaline cartilage. In some buildings similar to interver tebral discs, matrix composition and cell sorts range from one location to another, reflecting varying mechanical properties. Regions of fibrocartilage that are loaded predominantly in rigidity consist of huge crimped fibres of collagen type I embedded in a hydrated proteoglycan gel.
Premature fusion of sutures through the early progress section of the skull will end in numerous abnormalities hiv infection of macrophages generic prograf 0.5mg mastercard. The bones forming the base of the skull develop mainly through endochondral ossification mcgraw hill hiv infection cycle works prograf 0.5mg amex, and also play an necessary half within the general development of the face and the neurocranium hiv infection and seizures prograf 0.5mg low price. One of the most important is the spheno-occipital synchondrosis, between the body of the sphenoid anteriorly and the basilar a part of the occipital bone posteriorly; fusion between them is mostly completed between thirteen and 18 years of age. There are only two sites of synovial articulation related to the outside of the cranium, specifically: the temporomandibular joint and the atlanto-occipital joint (between the condyles of the occipital bone and the superior articular aspects of the atlas). Many essential neurological and vascular constructions enter and exit the cranium via foramina (openings). The skull is a prime web site for fractures resulting from trauma, which signifies that these constructions are vulnerable to damage following head harm. In addition to the principle foramina, irregular emissary foramina allow veins located externally on the face and scalp to communicate with those mendacity intracranially; unfold of infection alongside these routes might have critical scientific consequences. In the account of the cranium that follows, only generalized normal views will be thought-about. A more detailed account of every individual bone will be discovered related to the relevant regional text (Berkovitz and Moxham 1994). The higher part is formed by the frontal bone, which underlies the brow region above the orbits. Superomedial to each orbit is a rounded superciliary arch (more pronounced in males), between which there could additionally be a median elevation, the glabella. The glabella might show the stays of the interfrontal (metopic) suture, which often closes in the first postnatal yr (Weinzweig et al 2003), however persists in a small share of grownup skulls in various ethnic teams. The frontal bone articulates with the two nasal bones at the frontonasal sutures, marked by a depression at the root of the nostril; the point at which the frontonasal and internasal sutures meet is the anthropometric landmark known as the nasion. The upper, supraorbital, margin is fashioned completely by the frontal bone, interrupted on the junction of its sharp lateral two-thirds and rounded medial third by the supraorbital notch or foramen, which transmits the supraorbital vessels and nerve. The lateral margin of the orbit is fashioned largely by the frontal process of the zygomatic bone and is completed above by the zygomatic process of the frontal bone; the suture between them (frontozygomatic) lies in a palpable despair. The infraorbital margin is fashioned by the zygomatic bone laterally and the maxilla medially. The medial margin of the orbit is formed above by the frontal bone and under by the lacrimal crest of the frontal process of the maxilla. The central a half of the face is occupied primarily by the paired maxillae, separated by the anterior nasal aperture. Each maxilla contributes to the upper jaw, the ground and medial wall of the orbital cavity, the lateral wall of the nostril, the floor of the nasal aperture and the bone of the cheek. The anterior nasal backbone marks the upper limit of the intermaxillary suture at the lower margin of the anterior nasal aperture and is palpable in the nasal septum. Key: 1, sagittal suture; 2, frontal sinus; three, crista galli; four, mastoid air cells; 5, nasal septum; 6, inferior concha; 7, dens of axis. The short, thick zygomatic strategy of the maxilla has an indirect upper floor that articulates with the zygomatic bone at the zygomaticomaxillary suture. The frontal means of the maxilla ascends posterolateral to the nasal bone to articulate with it medially, the frontal bone superiorly and the lacrimal bone laterally. The anterior nasal aperture is piriform in shape, wider under than above and bounded by the paired nasal bones and maxillae. The upper boundary of the aperture is shaped by the nasal bones whereas the rest is fashioned by the maxillae. In life, several cartilages (septal, lateral nasal, main and minor alar) assist to delineate two nasal cavities. The form of these bones can be used fairly successfully to predict the form of the cartilaginous nostril in forensic facial reconstructions (Wilkinson and Rynn 2012). The decrease part of the face, below the nostril, is fashioned from the alveolar arch of the maxillae and the higher dentition, the physique of the mandible, the alveolar means of the mandible and the lower dentition. In the midline, the mental protuberance produces the characteristic prominence of the chin. The mental foramen, which transmits the psychological nerve and accompanying vessels, lies in the same vertical plane as the supraorbital and infraorbital foramina. Anteroposterior radiographs of the skull clearly present the central location of the paranasal air sinuses within the frontal bone, maxilla and ethmoid. These could be notably useful indicators of identity when postmortem pictures are compared with antemortem medical movies. The exterior occipital protuberance is a midline elevation on the occipital bone that may become significantly properly developed and palpable in males. Superior nuchal traces prolong laterally from the protuberance and characterize the boundary between the scalp and the neck. Inferior nuchal traces run parallel to , and below, the superior nuchal strains; a set of highest nuchal lines may sometimes happen above the superior traces. The exterior occipital protuberance, nuchal traces and roughened exterior floor of the occipital bone between the nuchal lines all afford attachment to muscles of the neck. The squamous a half of the frontal bone is anterior, the squamous a half of the occipital bone is posterior and the 2 parietal bones meet within the midline and separate the frontal from the occipital bone. The superior and inferior temporal lines run near the parietal eminence but are finest seen in a lateral view. The coronal suture marks the articulation between the posterior margin of the frontal bone and the anterior margins of the two parietal bones. It descends throughout the calvaria and initiatives inferiorly until it meets the junction between the greater wing of the sphenoid and the squamous temporal bone on the pterion. The sagittal suture runs in the midline between the two parietal bones and extends from the bregma anteriorly to the lambda posteriorly. The lambdoid suture delineates the articulation between the posterior borders of the right and left parietal bones and the superior border of the occipital bone. This diamond-shaped, membrane-filled area positioned between the 2 frontal and two parietal bones of the developing fetal cranium persists until roughly 18 months after birth. The lambda, at the junction of the sagittal and lambdoid sutures, represents the site of the posterior fontanelle, which persists for the primary 2�3 months after birth. A parietal foramen might pierce either or both parietal bones near the sagittal suture about 3. The vertex is the best point on the cranium and it often occupies a place within the center third of the sagittal suture. The superolateral region is occupied by the parietal bones, the mastoid area of the temporal bones makes up the inferolateral areas, and the central portion is occupied by the occipital bone, which is the reason why this side is also referred to as the occipital view. The parietal bones articulate with the occipital bone on the lambdoid suture, which extends inferiorly into the occipitomastoid and the parietomastoid sutures behind and above the mastoid processes, respectively. The lambda is the anthropometric point where the occipital bone meets the two parietal bones. Sagittal suture Parietal foramen Parietal foramen Lambda Lambdoid suture Parietal bone Interparietal bone Sutural bones Squamous suture Parietomastoid suture Occipitomastoid suture Mastoid foramen Mastoid notch Styloid process External occipital crest Mastoid process Superior nuchal line Interior nuchal line Inion External occipital protuberance (Transverse occipital suture, var. The temporal area may be divided into an upper temporal fossa and a decrease infratemporal fossa, separated by the position of the zygomatic arch. The upper temporal fossa is bounded inferiorly by the zygomatic arch, superiorly and posteriorly by the temporal traces, and anteriorly by the frontal process of the zygomatic bone, and is continuous inferiorly with the infratemporal fossa deep to the zygomatic arch. The temporal traces typically current anteriorly as distinct ridges but become much less distinguished as they arch posteriorly across the parietal bone. The inferior temporal line becomes extra outstanding as it curves down the posterior part of the squamous temporal bone, forming a supramastoid crest on the base of the mastoid process. The superior temporal line gives attachment to the temporal fascia whereas the inferior temporal line offers attachment for temporalis. The ground of the temporal fossa is formed by the frontal and parietal bones superiorly and the higher wing of the sphenoid and squamous part of the temporal bone inferiorly. All 4 bones of 1 facet meet at a roughly H-shaped sutural junction termed the pterion. This is an important anthropometric landmark as a end result of it commonly overlies both the anterior department of the middle meningeal artery and the lateral fissure of the cerebral hemisphere (Ma et al 2012). The pterion corresponds to the positioning of the anterolateral (sphenoidal) fontanelle of the neonatal skull, which closes in the third month after delivery. The vertical suture between the sphenoid and temporal bones, the sphenosquamosal suture, is fashioned by articulation between the posterior border of the higher wing of the sphenoid and the anterior border of the squamous part of the temporal bone. The lateral floor of the ramus of the mandible might be described briefly right here as a end result of it lies within the middle area of this view of the cranium.
Horresh I acute hiv infection how long does it last prograf 5mg buy visa, Poliak S antiviral resistance mechanisms discount prograf 0.5 mg line, Grant S et al 2008 Multiple molecular interactions determine the clustering of Caspr2 and Kv1 channels in myelinated axons antiviral vitamin c cheap prograf 0.5mg visa. A basic however complete neuroscience textbook Kempermann G 2011 Adult Neurogenesis 2. Matyash V, Kettenmann H 2010 Heterogeneity in astrocyte morphology and physiology. Murakoshi H, Yasuda R 2012 Postsynaptic signaling during plasticity of dendritic spines. Neher E, Sakaba T 2008 Multiple roles of calcium ions in the regulation of neurotransmitter launch. Perea G, Navarrete M, Araque A 2009 Tripartite synapses: astrocytes course of and management synaptic info. Poliak S, Peles E 2003 the local differentiation of myelinated axons at nodes of Ranvier. Robel S, Berninger B, Goetz, M 2011 the stem cell potential of glia: classes from reactive gliosis. Schmieg N, Memendez G, Schiavo G et al 2014 Signalling endosomes in axonal transport: journey updates on the molecular freeway. Segal V, Vlachos A, Korkotian E 2010 the backbone apparatus, synaptopodin, and dendritic spine plasticity. Seifert G, Schilling K, Steinh�user C 2006 Astrocyte dysfunction in neurological issues: a molecular perspective. Zujovic V, Thibaud J, Bachelin C et al 2011 Boundary cap cells are peripheral nervous system stem cells that might be redirected into central nervous system lineages. The latter embody neutrophil, eosinophil and basophil granulocytes, B lymphocytes and monocytes. These cells all contribute to the immune system of the human (for an overview of the immune system, see Murphy (2011)). Platelets are produced within the bone marrow as mobile fragments of megakaryocytes. Only erythrocytes and platelets are generally confined to the blood vascular system, whereas all leukocytes can leave the circulation and enter extravascular tissues. The numbers of cells doing so will increase significantly throughout inflammation brought on by native infections or tissue harm. The lymphoid tissues are the thymus, lymph nodes, spleen and the lymphoid follicles associated mainly with the alimentary and respiratory tracts. Lymphoid tissue also incorporates supportive stromal cells, that are non-haemopoietic in origin. Blood is an opaque fluid with a viscosity greater than that of water (mean relative viscosity four. It is brilliant red when oxygenated, in the systemic arteries, and darkish purple to purple when deoxygenated, in systemic veins. Plasma is a clear, yellowish fluid that incorporates many substances in answer or suspension: low-molecular-weight solutes give a imply freezing-point despair of zero. Plasma accommodates high concentrations of sodium and chloride ions, potassium, calcium, magnesium, phosphate, bicarbonate, traces of many other ions, glucose, amino acids and vitamins. The plasma is involved in the transport of most molecules which are launched or secreted by cells in response to pathological or physiological stimuli and so the routine chemical analysis of plasma is of great diagnostic importance. There is rising interest in utilizing metabolomics approaches for the highthroughput evaluation of small molecules or metabolites within the serum, as a possible aid to prognosis and understanding of disease (Psychogios et al 2011). Heparin can also be broadly used as an anticlotting agent as a outcome of it interferes with fibrin clot formation. Polycythaemia (increased red cell mass) can happen in individuals living at excessive altitude, or pathologically in conditions resulting in arterial hypoxia. Reduction in pink cell mass (anaemia) has many underlying causes but in uncommon cases could be as a result of structural defects in erythrocytes (see below). The erythrocyte biconcave disc is typically electrondense and almost fills the capillary lumen. The properties of their cell coat cause them to adhere to each other by their rims to form unfastened piles of cells (rouleaux). In normal blood, a quantity of cells assume a shrunken, star-like, crenated kind; this form can be reproduced by placing regular biconcave erythrocytes in a hypertonic resolution, which causes osmotic shrinkage. In hypotonic options erythrocytes take up water and turn out to be spherical; they might finally lyse to launch their haemoglobin (haemolysis), leaving red-cell ghosts. Erythrocytes have a plasma membrane that encloses primarily a single protein, haemoglobin, as a 33% solution. The plasma membrane of erythrocytes is 60% lipid and glycolipid, and 40% protein and glycoprotein. Glycophorins A and B (each with a molecular mass of roughly 50 kDa) span the membrane, and their negatively charged carbohydrate chains project from the outer floor of the cell. The filamentous protein, spectrin, is answerable for maintaining the form of the erythrocyte. A dimer is fashioned of 1 and 1 spectrin monomers, and two dimers then come together to form a tetramer (Machnicka et al 2013). These are joined by junctional complexes that contain (among different proteins) ankyrin, brief actin filaments, tropomyosin and protein four. This construction gives the membrane great flexibility; purple cells are deformable however regain their biconcave form and dimensions after passing through the smallest capillaries, which are 4 �m in diameter (Mohandas and Gallagher 2008). Erythrocyte membrane flexibility additionally contributes to the usually low viscosity of blood. Molecular defects in the cytoskeleton end in abnormalities of purple cell shape, membrane fragility, premature destruction of erythrocytes within the spleen and haemolytic anaemia (Iolascon et al 1998). They can work together with naturally occurring or induced antibodies in the plasma of recipients of an unmatched transfusion, causing agglutination and lysis of the erythrocytes. Erythrocytes of a single individual carry several various kinds of antigen, each sort belonging to an antigenic system by which a number of different antigens are attainable in different individuals. Leukocytes additionally bear highly polymorphic antigens encoded by allelic gene variants. In apply, leukocytes are often divided into two primary teams: specifically, those with prominent stainable cytoplasmic granules, the granulocytes, and those without. Granulocytes this group consists of eosinophil granulocytes, with granules that bind acidic dyes similar to eosin; basophil granulocytes, with granules that bind basic dyes strongly; and neutrophil granulocytes, with granules that stain solely weakly with both sort of dye. Haemoglobin Haemoglobin (Hb) is a globular protein with a molecular mass of 67 kDa. It consists of globulin molecules certain to haem, an ironcontaining porphyrin group. The oxygen-binding energy of haemoglobin is supplied by the iron atoms of the haem groups, and these are maintained within the ferrous (Fe++) state by the presence of glutathione throughout the erythrocyte. The haemoglobin molecule is a tetramer, made up of 4 subunits, each a coiled polypeptide chain holding a single haem group. Mutations in the haemoglobin chains can lead to a variety of pathologies (Forget and Bunn 2013). Neutrophil granulocytes Neutrophil granulocytes (neutrophils) are also referred to as polymorphonuclear leukocytes (polymorphs) because of their irregularly Lifespan Erythrocytes last between 100 and 120 days before being destroyed. As erythrocytes age, they become more and more fragile, and their floor charges decrease as their content of negatively charged membrane glycoproteins diminishes. Aged erythrocytes are taken up by the macrophages of the spleen (Mebius and Kraal 2005) and liver sinusoids, normally with out prior lysis, and are hydrolysed in phagocytic vacuoles the place the haemoglobin is split into its globulin and porphyrin moieties. Globulin is additional degraded to amino acids, which move into the general amino-acid pool. Iron is faraway from the porphyrin ring and either transported within the circulation certain to transferrin and used within the synthesis of latest haemoglobin within the bone marrow, or stored in the liver as ferritin or haemosiderin. The remainder of the haem group is converted within the liver to bilirubin and excreted in the bile. The recognition of effete erythrocytes by macrophages seems to take place by numerous mechanisms (Bratosin et al 1998). The neutrophil nucleus is extra segmented (four lobes are visible) and the granules are smaller and more electron-dense than within the basophil. Each haemoglobin molecule incorporates two -chains and two others, so that a number of mixtures, and hence a number of various varieties of haemoglobin molecule, are potential.
All but the seventh are also characterised by a foramen in each transverse course of antiviral zoster proven 5 mg prograf, the foramen transversarium (these foramina could also be absent or typically duplicated in the seventh cervical vertebra) xl 3 vr antiviral 0.5 mg prograf discount with amex. The first (atlas) antiviral tablets 0.5mg prograf visa, second (axis) and seventh (vertebra prominens) cervical vertebrae are atypical. It contains a variable amount of adipose tissue and platysma but is hardly demonstrable as a separate layer. The deep cervical fascia is conventionally subdivided into three sheets (superficial investing, center and deep layers) that surround the muscle tissue and viscera of the neck to various levels, and the carotid sheath, a condensation of deep fascia around the frequent and inner carotid arteries, internal jugular vein, vagus and ansa cervicalis. In health, the tissues within these spaces are either carefully utilized to each other or are full of comparatively free connective tissue. However, they provide potential routes by which unchecked infection might unfold throughout the head and neck, and between the tissue spaces of the face and the mediastinum. It is suspended from the styloid processes by the stylohyoid ligaments and gives attachment to the suprahyoid and infrahyoid teams of muscle tissue. The skeletal framework of the larynx is fashioned by a sequence of cartilages interconnected by ligaments and fibrous membranes, and moved by a variety of muscles (Ch. The laryngeal cartilages are the single cricoid, thyroid and epiglottic cartilages, and the paired arytenoid, cuneiform, corniculate and tritiate cartilages. It may be divided into the cranium, consisting of the calvaria (brain box) and basicranium, which collectively encompass and shield the mind; a delicate facial skeleton composed primarily of thin-walled bones, some of which include air-filled cavities which may be known collectively as the paranasal sinuses; and the mandible. The cranial cavity accommodates the brain and the intracranial parts of the cranial nerves; the blood vessels that offer and drain the mind and the haemopoietic marrow of the overlying bones; the meninges (dura, arachnoid and pia mater); and the cerebrospinal fluid within the subarachnoid house. The cavity is incompletely divided by dural partitions, notably the falx cerebri, mendacity between the cerebral hemispheres, and the tentorium cerebelli, lying between the cerebellum and occipital lobes. Almost all of the venous blood from the brain and cranial bones drains through sinuses lying between the endosteal and meningeal layers of the dura mater into the inner jugular vein. The superior tarsal muscle, sphincter and dilator pupillae and the ciliary muscle are composed of clean muscle (Chs forty one and 42). Subclavianartery the subclavian arteries give off a quantity of branches that supply constructions in the head and neck. The vertebral arteries supply the higher spinal twine, brainstem, cerebellum and occipital lobe of the cerebrum. They cross by way of the foramina transversaria of the primary six cervical vertebrae, enter the cranial cavity through the foramen magnum and unite at the lower border of the pons to type the basilar artery (hence this method is often referred to as the vertebrobasilar system). Branches from the thyrocervical trunk provide the inferior poles of the thyroid gland and the parathyroid glands, the larynx and the pharynx, and branches from the costocervical trunks provide deep cervical muscular tissues. Carotidsystem the cervical portion of the common carotid artery is analogous on each side. Each lies inside the carotid sheath of deep cervical fascia, together with the internal jugular vein and vagus nerve. At the level of the higher border of the thyroid cartilage (C4), the common carotid artery bifurcates into exterior and inside carotid arteries (see below). The exterior carotid artery passes upwards on both aspect of the neck, inclined at first barely forwards after which backwards and slightly laterally. It normally gives off the ascending pharyngeal, superior thyroid, lingual, facial, occipital and posterior auricular arteries, and then enters the parotid salivary gland the place it divides into its terminal branches, the superficial temporal and maxillary arteries. The branches of the exterior carotid artery supply the face, scalp, tongue, higher and lower teeth and gingivae, palatine tonsil, paranasal sinuses and nasopharyngeal tube, external and middle ears, pharynx, larynx and superior pole of the thyroid gland. They also anastomose with branches of the internal carotid arteries on the scalp, forehead and face, in the orbit, nasopharynx and nasal cavity, and with branches of the subclavian artery within the pharynx, larynx and thyroid glands. The inside carotid artery supplies most of the ipsilateral cerebral hemisphere, eye and accessory organs, the forehead and, partially, the external nostril, nasal cavity and paranasal sinuses. It passes up the neck anterior to the transverse processes of the higher three cervical vertebrae and enters the cranial cavity via the carotid canal in the petrous a half of the temporal bone. The artery has no branches within the neck and so is well distinguishable from the external carotid artery, should the latter require ligation. Superficial veins in the end drain into either the exterior, anterior or posterior exterior jugular veins; they drain a a lot smaller volume of tissue than the deep veins. Deep veins are inclined to drain into either the internal jugular vein or the subclavian vein. The inner jugular vein drains blood from the skull, brain, superficial face and much of the neck. At its junction with the inner jugular vein, the left subclavian vein usually receives the thoracic duct, and the best subclavian vein receives the proper lymphatic duct. The vertical chain consists of superior and inferior teams of nodes associated to the carotid sheath. All lymph vessels of the pinnacle and neck drain into the deep cervical nodes, either instantly from the tissues or indirectly by way of nodes in outlying teams. Lymph is returned to the systemic venous circulation via both the proper lymphatic duct or the thoracic duct. Reflexes A number of reflexes involving buildings in the head and neck are mediated by sensory and motor branches of sure of the cranial nerves, coordinated by way of applicable nuclei in the brainstem. They are individually named and numbered (using Roman numerals) in a rostrocaudal sequence (see Table 16. Some are functionally blended, others are both purely motor or purely sensory, and a few additionally carry pre- or postganglionic parasympathetic fibres which may be secretomotor to the salivary and lacrimal glands or motor to the smooth muscle inside the eyeball and orbit. Branches of the oculomotor, trochlear, trigeminal, abducens, facial, glossopharyngeal, vagus, accent and hypoglossal nerves supply muscle groups inside the eyeball, face, neck, pharynx, larynx and tongue. Branches of the trigeminal, glossopharyngeal and vagus nerves transmit common sensory information from the skin of the face and part of the scalp; the epithelium lining the oral and nasal cavities, the paranasal sinuses, middle ear, pharynx and larynx, and the dorsal surface of the tongue and the cornea; the intracranial meninges; and the periosteum and bones of the cranium. The olfactory, optic, trigeminal, facial, vestibulocochlear and vagus nerves contain axons that transmit the particular sensations of olfaction, imaginative and prescient, hearing, steadiness and style. The olfactory nerve is the only sensory cranial nerve that projects on to the cerebral cortex quite than not directly via the thalamus (Ch. The other ten pairs of cranial nerves are hooked up to the brainstem or, within the case of the accent nerve, to the higher cervical spinal twine; their component fibres come up from or terminate in named cranial nerve nuclei. The cranial nerves pass by way of named foramina within the cranium, often with named vessels. Cutaneous branches of the dorsal rami of the second, third, fourth and fifth cervical nerves innervate the scalp and the skin over the again of the neck, and motor branches of the entire cervical dorsal rami provide cervical postvertebral muscular tissues. All of the cervical ventral rami provide anterior and lateral teams of prevertebral muscular tissues. The higher four cervical ventral rami kind the cervical plexus, whose branches collectively innervate the infrahyoid strap muscular tissues and the diaphragm, and the skin masking the lateral and anterior parts of the neck, and the angle of the mandible. The lower 4 cervical ventral rami, together with most of the first thoracic ventral ramus, form the brachial plexus. Some preganglionic fibres could synapse around cells in the hilum of the submandibular gland. Stimulation of the chorda tympani dilates the arterioles in both glands, in addition to having a direct secretomotor effect. Pterygopalatineganglion Preganglionic axons travel within the greater petrosal department of the facial nerve and the nerve of the pterygoid canal, and relay in the pterygopalatine ganglion. Postganglionic secretomotor axons innervate secretory acini and blood vessels in the palatine, pharyngeal and nasal mucosa through the palatine and nasal nerves, but whether additionally they innervate the lacrimal gland via the zygomatic and zygomaticotemporal branches of the maxillary nerve, as was once thought, is much less sure. It is likely that postganglionic orbital branches, carrying a mix of postganglionic parasympathetic and somatic sensory axons, cross via the inferior orbital fissure and innervate the lacrimal gland and ophthalmic artery immediately. Some axons pass into the cranial cavity via the ethmoidal vessels to innervate the choroid; the pterygopalatine ganglion is believed to be the main source of parasympathetic input to the choroid. Ciliaryganglion Preganglionic axons originate in the Edinger�Westphal preganglionic nucleus of the midbrain (Ch. They journey through a branch of the oculomotor nerve (nerve to the inferior oblique) to the ciliary ganglion, the place they synapse. Postganglionic fibres journey within the quick ciliary nerves, which pierce the scleral coat of the eyeball and run forwards in the perichoroidal house to enter the ciliary muscle and sphincter pupillae. Their activation mediates lodging of the eye to near objects and pupillary constriction.






